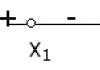
Эмбриональный период имеет следующие этапы: дробление, гаструляция, гисто- и органогенез.
. Дробление. В результате дробления образуется бластула;
. Гаструляция. В результате гаструляции образуются зародышевые листки;
. Гисто- и органогенез - образование тканей и осевых органов зародыша.
При личиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из яй-
цевых оболочек. При неличиночной форме онтогенеза этот период начинается с образования зиготы и заканчивается выходом из зародышевых оболочек. При внутриутробной форме онтогенеза - с образования зиготы до рождения особи.
Дробление
В результате оплодотворения образуется зигота, которая начинает дробиться. Дробление сопровождается митотическими делениями. Между делениями в короткой интерфазе отсутствует пост- митотический период, а синтез ДНК начинается в телофазе предыдущего митотического деления. Не происходит роста зародыша. Суммарный объем зародыша не изменяется. Клетки, образующиеся в процессе дробления, называются бластомерами, а зародыш - бластулой. Тип дробления зависит от количества и распределения желтка в яйцеклетке (рис. 67).
Рис. 67. Типы яиц и способы их дробления (яйца и зародыши изображены вверх анимальным полюсом).
I - изолецитальное яйцо (а) и его равномерное дробление (б); II и III - телолецитальные яйца (в, д), и их дробления - дискоидальное (г) и неравномерное (е); IV - центролецитальное яйцо (ж) и его поверхностное дробление (з).
Типы дробления
Дробление может быть полным равномерным, полным неравномерным, неполным дискоидальным, поверхностным (рис. 68).
Полное равномерное дробление характерно для изолецитальных яиц - например для ланцетника. Ядро зиготы делится митозом на

Рис. 68. Типы яиц и соответствующие им типы дробления.
два, затем делится цитоплазма. Борозда дробления проходит по меридиану, образуются два бластомера. Затем снова делится ядро и на поверхности зародыша появляется вторая борозда дробления, идущая по меридиану, перпендикулярно первой. Образуются 4 бластомера, 3-я борозда проходит по экватору и делит его на 8 частей. Затем происходит чередование меридионального и экваториального дроблений. Число бластомеров увеличивается. Зародыш на стадии 32 бластомеров называется морулой. Дробление продолжается до образования зародыша, похожего на пузырек, стенки которого образованы одним слоем клеток, называемых бластодермой. Бластомеры расходятся от центра зародыша, образуя полость, которая называется первичной, или бластоцелью. Бластомеры имеют одинаковый размер. В результате такого дробления образуется целобластула.
Полное неравномерное дробление свойственно телолецитальным яйцам с умеренным содержанием желтка, например у лягушек. 1-я и 2-я борозды дробления проходят по меридианам и полностью делят яйцо на 4 части, 3-я борозда смещена в сторону анимального полюса, где нет желтка. Бластомеры имеют неодинаковую величину: на анимальном полюсе они меньше (микромеры), на вегетативном больше (макромеры). Желток затрудняет дробление, и поэтому дробление макромеров идет медленнее, чем микромеров. Стенка бластулы состоит из нескольких рядов клеток. Первичная полость мала и смещена к анимальному полюсу. Образуется амфибластула.
Неполному дискоидальному дроблению подвергаются телолецитальные яйца с большим содержанием желтка, например у рептилий, птиц. Дробление идет только на анимальном полюсе. 1-я и 2-я борозды дробления проходят по меридиану перпендикулярно друг другу, 3-я борозда смещена к анимальному полюсу. В результате этого образуется зародышевый диск. Бластоцель располагается под слоем бластодермы в виде щели. Бластула называется дискобластулой.
Неполное поверхностное дробление характерно для центролецитальных яиц, например для членистоногих. Ядра центролециталь-
ных яиц многократно делятся и перемещаются к периферии, где в цитоплазме нет желтка. Цитоплазма образует бластомеры. Бласту- ла имеет один слой бластомеров. Бластоцель заполнена желтком. Такая бластула называется перибластулой.
Гаструляция
По окончании периода дробления у многоклеточных животных наступает период образования зародышевых листков - гаст- руляция. Гаструляция связана с перемещением эмбрионального материала. Сначала образуется ранняя гаструла, имеющая два зародышевых листка (эктодерму и энтодерму) , затем поздняя гаструла, когда формируется 3-й зародышевый листок - мезодерма. Зародыш, образовавшийся в результате гаструляции, называется гаструлой.
Образование ранней гаструлы происходит следующими способами:
. иммиграцией (выселением клеток), у кишечнополостных;
. инвагинацией (впячиванием), у ланцетника;
. эпиболией (обрастанием), у лягушки;
. деляминацией (расщеплением), у кишечнополостных.
При иммиграции часть клеток бластодермы с поверхности зародыша уходит в бластоцель. Образуется наружный слой - эктодерма и внутренний - энтодерма. Бластоцель заполнена клетками.
При инвагинации определенный участок бластодермы (вегетативный полюс) прогибается внутрь и достигает анимального полюса. Образуется двуслойный зародыш-гаструла. Наружный слой клеток - эктодерма, внутренний - энтодерма выстилает полость первичной кишки (гастроцель). Отверстие, при помощи которого полость сообщается с внешней средой, называется первичным ртом - бластопором. У первичноротых животных (черви, моллюски, членистоногие) он превращается в ротовое отверстие, у вторичноро- тых - в анальное отверстие, а рот образуется на противоположном конце (хордовые).
Эпиболия характерна для животных, которые развиваются из телолецитальных яиц. Образование гаструлы идет за счет быст-
рого размножения микромеров, которые обрастают вегетативный полюс. Макромеры оказываются внутри зародыша. Образование бластопора не происходит и нет гастроцели.
Деляминация встречается у кишечнополостных, бластула которых похожа на морулу. Клетки бластодермы делятся на наружный и внутренний слои. Наружный слой образует эктодерму, внутренний - энтодерму (рис. 69).
У всех многоклеточных, кроме губок и кишечнополостных, возникает 3-й зародышевый листок - мезодерма. Формирование мезодермы происходит двумя способами: телобластическим или энтероцельным.
Телобластический способ свойствен первичноротым. На границе между эктодермой и энтодермой по бокам от бластопора

Рис. 69. Типы гаструл.
а - инвагинационная гаструла; б, в - две стадии развития иммиграционной гаструлы; г, д - две стадии развития деляминационной гаструлы; е, ж - две стадии развития эпиболической гаструлы; 1 - эктодерма; 2 - энтодерма; 3 - бластоцель.
клетки - телобласты - начинают делиться и дают начало мезодерме.
Энтероцельный способ характерен для вторичноротых. Клетки, формирующие мезодерму, обособляются в виде карманов первичной кишки. Полости карманов превращаются в целом. Мезодерма делится на отдельные участки - сомиты, из которых образуются определенные ткани и органы.
Гисто- и органогенез
После образования мезодермы начинается процесс гисто- и органогенеза. Сначала формируются осевые органы - нервная трубка, хорда, затем все остальные органы (рис. 70).
У ланцетника из эктодермы на спинной стороне зародыша образуется нервная трубка. Остальная эктодерма формирует кожный эпителий и его производные. Из энто- и мезодермы под нервной трубкой формируется хорда. Под хордой находится кишечная трубка, по бокам от хорды - мезодерма сомитов. Наружная часть сомита, прилегающая к эктодерме, называется дермотомом. Из нее формируется соединительная ткань кожи. Внутренняя часть - склеротом - дает начало скелету. Между дермотомом и склеротомом находится миотом, дающий начало поперечно-полосатой мускулатуре. Под сомитами находятся ножки (нефрогонотом), из которых формируется мочеполовая система.
Целомические мешки формируются симметрично по бокам. Стенки целомических мешков, обращенные в сторону кишечника, на-

Рис. 70. Образование мезодермы (по Шимкевичу, 1925, модифицировано).
а - у первичноротых; б - у вторичноротых;
1 - эктодерма; 2 - мезенхима; 3 - энтодерма; 4 - телобласт (а) и целомическая мезодерма (б).
зывают спланхноплеврой, в сторону эктодермы - соматоплеврой. Эти листки участвуют в образовании сердечно-сосудистой системы, плевры, брюшины, перикарда (рис. 71).
Таким образом, из эктодермы формируются кожный эпителий, кожные железы, эмаль зубов, волосы, ногти, когти.
Из энтодермы образуются эпителий средней кишки, печень, поджелудочная железа, щитовидная железа, тимус, эпителий дыхательной системы.
Мезодерма участвует в образовании мышц, соединительной ткани, костной ткани, каналов выделительной системы, кровеносной системы, части ткани половых желез.
Сущность стадии дробления. Дробление - это ряд последовательных митотических делений зиготы и далее бластомеров, заканчивающихся образованием многоклеточного зародыша - бластулы. Первое деление дробления начинается после объединения наследственного материала пронуклеусов и образования общей метафазной пластинки. Возникающие при дроблении клетки называют бластомерами (от греч. бласте- росток, зачаток). Особенностью митотических делений дробления является то, что с каждым делением клетки становятся все мельче и мельче, пока не достигнут обычного для соматических клеток соотношения объемов ядра и цитоплазмы. У морского ежа, например, для этого требуется шесть делений и зародыш состоит из 64 клеток. Между очередными делениями не происходит роста клеток, но обязательно синтезируется ДНК.
Все предшественники ДНК и необходимые ферменты накоплены в процессе овогенеза. В результате митотические циклы укорочены и деления следуют друг за другом значительно быстрее, чем в обычных соматических клетках. Сначала бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость - бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, образуя стенку бластулы - бластодерму. Общий размер зародыша к концу дробления на стадии бластулы не превышает размера зиготы.
Главным результатом периода дробления является превращение зиготы в многоклеточный односменный зародыш.
Морфология дробления. Как правило, бластомеры располагаются в строгом порядке друг относительно друга и полярной оси яйца. Порядок, или способ, дробления зависит от количества, плотности и характера распределения желтка в яйце. По правилам Сакса - Гертвига клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы, а веретено клеточного деления - в направлении наибольшей протяженности этой зоны.
В олиго- и мезолецитальных яйцах дробление полное, или голобластическое. Такой тип дробления встречается у миног, некоторых рыб, всех амфибий, а также у сумчатых и плацентарных млекопитающих. При полном дроблении плоскость первого деления соответствует плоскости двусторонней симметрии. Плоскость второго деления проходит перпендикулярно плоскости первого. Обе борозды первых двух делений меридианные, т.е. начинаются на анимальном полюсе и распространяются к вегетативному полюсу. Яйцевая клетка оказывается разделенной на четыре более или менее равных по размеру бластомера. Плоскость третьего деления проходит перпендикулярно первым двум в широтном направлении. После этого в мезолецитальных яйцах на стадии восьми бластомеров проявляется неравномерность дробления. На анимальном полюсе четыре более мелких бластомера - микромеры, на вегетативном - четыре более крупных - макромеры. Затем деление опять идет в меридианных плоскостях, а потом опять в широтных.
В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или мероб-ластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным.
При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным. Оно типично для хордовых и иглокожих. В природе встречаются и другие варианты пространственного расположения бластомеров при дроблении, что определяет такие его типы, как спиральное у моллюсков, билатеральное у аскариды, анархичное у медузы.
Замечена зависимость между распределением желтка и степенью синхронности деления анимальных и вегетативных бластомеров. В олиголецитальных яйцах иглокожих дробление почти синхронное, в мезолецитальных яйцевых клетках синхронность нарушена после третьего деления, так как вегетативные бластомеры из-за большого количества желтка делятся медленнее. У форм с частичным дроблением деления с самого начала асинхронны и бластомеры, занимающие центральное положение, делятся быстрее.
Рис. 7.2. Дробление у хордовых животных с разным типом яйцеклетки.
А - ланцетник; Б - лягушка; В - птица; Г - млекопитающее:
I -два бластомера, II- четыре бластомера, III- восемь бластомеров, IV- морула, V- бластула;
1 -борозды дробления, 2 -бластомеры, 3- бластодерма, 4- бластоиель, 5- эпибласт, 6- гипобласт, 7-эмбриобласт, 8- трофобласт; размеры зародышей на рисунке не отражают истинных соотношений размеров

Рис. 7.2. Продолжение
К концу дробления образуется бластула. Тип бластулы зависит от типа дробления, а значит, от типа яйцеклетки. Некоторые типы дробления и бластул представлены на рис. 7.2 и схеме 7.1. Более подробное описание дробления у млекопитающих и человека см. разд. 7.6.1.
Особенности молекулярно-генетических и биохимических процессов при дроблении. Как было отмечено выше, митотические циклы в периоде дробления сильно укорочены, особенно в самом начале.
Например, весь цикл деления в яйцах морского ежа длится 30-40 мин при продолжительности S-фазы всего 15 мин. gi- и 02-периоды практически отсутствуют, так как в цитоплазме яйцевой клетки создан необходимый запас всех веществ, и тем больший, чем она крупнее. Перед каждым делением происходит синтез ДНК и гистонов.

Скорость продвижения репликационной вилки по ДНК в ходе дробления обычная. Вместе с тем в ДНК бластомеров наблюдается больше точек инициации, чем в соматических клетках. Синтез ДНК идет во всех репликонах одновременно, синхронно. Поэтому время репликации ДНК в ядре совпадает с временем удвоения одного, притом укороченного, репликона. Показано, что при удалении из зиготы ядра дробление происходит и зародыш доходит в своем развитии почти до стадии бластулы. Дальнейшее развитие прекращается.
В начале дробления другие виды ядерной активности, например транскрипция, практически отсутствуют. В разных типах яиц транскрипция генов и синтез РНК начинаются на разных стадиях. В тех случаях, когда в цитоплазме много различных веществ, как, например, у земноводных, транскрипция активируется не сразу. Синтез РНК у них начинается на стадии ранней бластулы. Напротив, у млекопитающих синтез РНК уже начинается на стадии двух бластомеров.
В периоде дробления образуются РНК и белки, аналогичные синтезируемым в процессе овогенеза. В основном это гистоны, белки клеточных мембран и ферменты, необходимые для деления клеток. Названные белки используются сразу же наравне с белками, запасенными ранее в цитоплазме яйцеклеток. Наряду с этим в период дробления возможен синтез белков, которых не было ранее. В пользу этого свидетельствуют данные о наличии региональных различий в синтезе РНК и белков между бластомерами. Иногда эти РНК и белки начинают действовать на более поздних стадиях.
Важную роль в дроблении играет деление цитоплазмы - цитотомия. Она имеет особое морфогенетическое значение, так как определяет тип дробления. В процессе цитотомии сначала образуется перетяжка с помощью сократимого кольца из микрофиламентов. Сборка этого кольца проходит под непосредственным влиянием полюсов митотического веретена. После цитотомии бластомеры олиголецитальных яиц остаются связанными между собой лишь тоненькими мостиками. Именно в это время их легче всего разделить. Это происходит потому, что цитотомия ведет к уменьшению зоны контакта между клетками из-за ограниченной площади поверхности мембран
Сразу после цитотомии начинается синтез новых участков клеточной поверхности, зона контакта увеличивается и бластомеры начинают плотно соприкасаться. Борозды дробления проходят по границам между отдельными участками овоплазмы, отражающим явление овоплазматической сегрегации. Поэтому цитоплазма разных бластомеров различается по химическому составу.
Дробление
I
Дробле́ние
в технике, процесс разрушения кусков твёрдого материала для уменьшения их размера. Куски разрушаются внешними силами, преодолевающими силы сцепления между частицами материала. Д. принципиально не отличается от измельчения (См. Измельчение). Условно считают, что при Д. получают продукты крупнее, а при измельчении мельче 5 мм
. Способы Д. (рис. 1
): раздавливание, раскалывание, истирание и удар. Прочные и абразивные материалы дробят преимущественно раздавливанием, прочные и вязкие - раздавливанием с истиранием, мягкие и хрупкие - раскалыванием и ударом. Работа Д. затрачивается на деформацию куска и на образование новой поверхности мелких кусков. Большая часть затраченной энергии рассеивается в виде тепла, и только небольшая доля преобразуется в свободную поверхностную энергию твёрдого тела. Полная работа Д. равна сумме работ на деформацию и на образование новых поверхностей. Эта обобщённая формула предложена П. А. Ребиндер ом (1944). Для приближённых расчётов принимают, что работа по Д. куска размером Д
при данной степени Д. прямо пропорциональна Д 2,5
. Д. характеризуют степенью Д., т. е. отношением размеров наибольших кусков в материале до и после Д. Др. показатель - удельный расход энергии, т. е. количество квт
·ч
на 1 т
дроблёного материала. Д. комбинируют, как правило, с Грохочение м. Различают Д. в открытом (рис. 2
, а) и замкнутом (рис. 2
, б) цикле. В 1-м случае готовый по крупности продукт отсеивают на грохоте перед дробилкой, а также получают после Д.; во 2-м - материал после дробилки просеивается на грохоте на крупный и мелкий (готовый); крупный материал возвращается для додрабливания в ту же дробилку.
Для получения высоких степеней Д. применяют последовательно несколько приёмов (стадий) Д. При обогащении руды дробят в 2, 3 или 4 стадии, удельный расход энергии на Д. от кусков размерами 900-1200 мм
до кусков 25 мм
- 1,5-3 квт
·ч
на 1 т
руды. Д. ручное и огневое было известно за 3000 лет до н. э. Простейшие машины - падающие песты (толчеи), приводимые в движение водяным колесом, применялись уже в средние века и описаны Г. Агриколой. Машинное Д. развивается с начала 19 в. (см. Дробилка). С 50-х гг. в СССР и др. странах исследуют гидровзрывные, термические, электротермические и др. способы Д., однако на ближайшие десятилетия главными останутся описанные механические способы. Д. применяют в горной, металлургической, химической, пищевой промышленности, в строительстве и сельском хозяйстве. Лит.:
Левенсон Л. Б., Клюев Г. М., Производство щебня, М., 1959; Андреев С. Е., Зверевич В. В., Перов В. А., Дробление, измельчение и грохочение полезных ископаемых, 2 изд., М., 1966; Труды Европейского совещания по измельчению, пер. с нем., М., 1966; Арш Э. И., Виторт Г. К., Черкасский Ф. Б., Новые методы дробления крепких горных пород, К., 1966; Пономарев И. В., Дробление и грохочение углей, М., 1970. В. А. Перов.
яйца, его сегментация, ряд последовательных делений яйца, в результате которых оно разделяется на всё более мелкие клетки (Бластомеры). Д. - непременная стадия развития всех многоклеточных животных. Обычно начинается после сближения мужского и женского Пронуклеус ов (см. Оплодотворение) и объединения их хромосом на веретене 1-го деления Д. У некоторых животных происходит Д. неоплодотворённых яиц (см. Партеногенез). Иногда оплодотворённые яйца находятся некоторое время в покое (см. Диапауза) и побуждаются к развитию изменением внешних условий (например, температуры окружающей среды). Вначале, в период синхронных делений, ядра во всех бластомерах делятся с одинаковым и постоянным ритмом, ядерный цикл короткий; у разных групп животных этот период неодинаков по продолжительности, а у млекопитающих отсутствует. Затем, в период асинхронных делений, или бластуляции (См. Бластуляция), ядерный цикл удлиняется, нарушается синхронность в делении разных ядер, на стадии интерфазы в них начинается синтез рибонуклеиновой кислоты (РНК), обнаруживается их морфогенетическая функция. Разделение цитоплазмы (цитотомия) следует за делением ядер (кариотомия), но, как правило, отстаёт от него. Д. не сопровождается ростом, и зародыш сохраняет исходные размеры яйца. По окончании Д. зародыш достигает стадии бластулы (См. Бластула). На характер Д. влияют количество и распределение желтка в цитоплазме яиц. Содержащие сравнительно мало равномерно распределённого желтка гомолецитальные яйца претерпевают полное равномерное Д. Чаще желток распределён в цитоплазме яйца неравномерно (телолецитальные и центролецитальные яйца). Область, содержащая больше желтка, делится медленнее бедной желтком - полное неравномерное Д., или совсем не делится - частичное Д. Яйца, претерпевающие полное Д., называют голобластическими, частичное Д., - меробластическими. К голобластическим относятся гомолецитальные (например, яйца многих беспозвоночных, ланцетника, млекопитающих) и часть телолецитальных (например, яйца некоторых членистоногих, большинства земноводных), которые претерпевают полное, но неравномерное Д. (мелкие бластомеры называются микромерами, средние - мезомерами, крупные - макромерами). К меробластическим яйцам относятся часть телолецитальных и центролецитальные с большим количеством желтка. В таких телолецитальных яйцах делится только бедная желтком анимальная часть яйца, которая последовательно разделяется на 2, 4 и большее число бластомеров, образующих диск клеток на поверхности недробящегося желтка, - дискоидальное Д. Оно характерно для яиц скорпионов, головоногих моллюсков, акуловых и костистых рыб, птиц, пресмыкающихся и низших млекопитающих. В результате дискоидального Д. образуется дискобластула, полость которой ограничена размерами бластодермы. Частичное Д. характерно и для центролецитальных яиц большинства членистоногих. После оплодотворения ядро начинает делиться. После нескольких синхронных делений ядра с окружающей их цитоплазмой перемещаются по цитоплазматическим мостикам в поверхностный слой цитоплазмы, который сначала представляет собой симпласт, затем вокруг каждого ядра обособляется отдельная клетка. В результате образуется зародыш, стенка которого состоит из одного слоя клеток (бластодермы), а центральная часть занята неразделившимся желтком с находящимися в нём клетками (вителлофагами); такой зародыш называется перибластулой, а Д. - поверхностным, или синцитиальным. На характер Д. влияют и свойства цитоплазмы яйца, определяющие положение веретён деления и, как следствие, положение бластомеров друг относительно друга, поскольку плоскость дробления всегда перпендикулярна оси веретена. По признаку относительного положения бластомеров при полном Д. различают радиальное, спиральное, билатеральное и двусимметричное Д. При радиальном Д., свойственном многим кишечно-полостным, иглокожим, земноводным и др., бластомеры располагаются так, что любая плоскость, которую можно провести через анимально-вегетативную ось яйца, является плоскостью симметрии. Первые 2 борозды проходят обычно меридионально, а 3-я - экваториально; затем происходит чередование меридиональных и экваториальных делений. В результате радиального Д. образуется многоклеточный пузырёк с полостью - целобластула. При спиральном Д., характерном для большинства турбеллярий, кольчецов, немертин, моллюсков и др., микромеры, отделяющиеся от первых 4 бластомеров (макромеров), располагаются в промежутках между ними. Происходят смещения бластомеров верхнего яруса относительно нижнего вправо - дексиотропное Д., или влево - леотропное Д. При спиральном Д. зародыш на стадии бластулы имеет полость (неравномерная целобластула) или не имеет её (стерробластула). При билатеральном Д. (у круглых червей, асцидий), а также на поздних стадиях спирального Д. деления происходят так, что у зародышей имеется только одна плоскость симметрии. Двусимметричное Д. наблюдается очень редко (гребневики) и характеризуется наличием двух плоскостей симметрии. См. схему строения яиц, типов их дробления и типов бластул.
Тот или иной тип Д. обычно присущ большинству представителей того или иного класса животных, но иногда в пределах класса наблюдаются разные типы Д. Так, среди земноводных, большинству которых свойственно полное неравномерное Д., у безногих земноводных имеется дискоидальное Д.; у млекопитающих происходит как дискоидальное (однопроходные), так и полное Д. (все высшие млекопитающие). Последнее по ряду признаков (обособление зародышевого диска и внезародышевой части) приближается к дискоидальному, от которого оно произошло. В результате полного Д. возникает бластоциста; часть её стенки, представленная плотным скоплением клеток, образует зародышевый диск, остальная часть представляет собой Трофобласт . В процессе Д. я́дра делятся равномерно (ядра всех бластомеров несут полный объём генетической информации и равноценны как друг другу, так и ядру зиготы), а цитоплазма делится неравномерно. Различия в свойствах цитоплазмы первых бластомеров у разных животных выражены в неодинаковой степени и зависят от уровня дифференцировки её в оогенезе (см. Сегрегация ооплазматическая). У одних животных при искусственном разделении двух первых бластомеров из каждого образуется целый зародыш, у других - только его часть, т.к. в яйцах разных животных к началу Д. цитоплазма достигает разной степени дифференцировки (См. Дифференцировка) (наиболее ранняя дифференцировка характерна для яиц со спиральным, билатеральным и поверхностным Д.). На этом основании иногда различают регуляционные и мозаичные яйца. В процессе Д. равноценные по генотипу ядра приходят во взаимодействие с качественно различающейся в разных бластомерах цитоплазмой, что является условием дифференциальной реализации в них генетической информации (см. Зародышевое развитие). Лит.:
Иванов П. П., Руководство по общей и сравнительной эмбриологии, Л., 1945; Токин Б. П., Общая эмбриология, , М., 1970. Т. А. Детлаф.
Схема строения яиц, типы их дробления и типы бластул: А - целобластула (1 - равномерная, 2 - неравномерная: а - бластоцель); Б - стерробластула; В - дискобластула (а - бластоцель, б - желток); Г - перибластула.
Большая советская энциклопедия. - М.: Советская энциклопедия . 1969-1978 .
Синонимы :Антонимы :
Смотреть что такое "Дробление" в других словарях:
См. разделение... Словарь русских синонимов и сходных по смыслу выражений. под. ред. Н. Абрамова, М.: Русские словари, 1999. дробление разбиение, разделение, раздел, деление, членение, расчленение; размежевание, литотрипсия, толочение,… … Словарь синонимов
- (a. breaking, crushing; н. Brechen, Zerkleinerung, Quetschen; ф. broyage, concassage; и. molienda) процесс разрушения кусков руды, угля и другого твёрдого материала с целью получения требуемой крупности (более 5 мм), гранулометрич.… … Геологическая энциклопедия
Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд… … Википедия
ДРОБЛЕНИЕ, дробления, мн. нет, ср. (книжн.). 1. Действие по гл. дробить и дробиться. Дробление камня. Дробление темы. 2. Процесс деления оплодотворенного яйца на отдельные клетки (биол.). Толковый словарь Ушакова. Д.Н. Ушаков. 1935 1940 … Толковый словарь Ушакова
Дробление - (помол, измельчение) – (керам.) уменьшение размеров материалов в зависимости от их твердости. [ГОСТ Р 54868 2011] Дробление огнеупорного сырья [неформованного огнеупора] – измельчение кусков огнеупорного сырья [неформованного… … Энциклопедия терминов, определений и пояснений строительных материалов
В технике процесс разрушения кусков твердого материала на более мелкие. В зависимости от крупности исходного материала различают: крупное (от 1000 до 100 мм), среднее (от 100 до 40 мм), мелкое (30 5 мм) дробление … Большой Энциклопедический словарь
Яйца, ряд последовательных митотич. делений оплодотворённого яйца, в результате к рых оно, не увеличиваясь в размерах, разделяется на всё более мелкие клетки бластомеры. Д. непременный период онтогенеза всех многоклеточных животных. Обычно… … Биологический энциклопедический словарь
Дробление
Дроблением называют ряд последовательных митотических делений зиготы, в результате которых она разделяется на всё более мелкие клетки - бластомеры. При этом рост новообразующихся клеток (бластомеров) крайне ограничен. Образуются 2, 4, 8, 16, 32 и т.д. бласто-
меров (рис. 95), а в конце концов возникает зародыш, состоящий из многих тысяч бластомеров, называемый бластулой. Бластула - пузырчатое образование, построенное из одного или нескольких слоёв клеток (бластодермы), окружающих полость - бластоцель (рис. 96). Яйцеклетка характеризуется анимально-вегетативным градиентом. Различают анимальный полюс, у которого происходило выделение редукционных телец, и вегетативный полюс, у которого скапливается желток при неравномерном распределении в цитоплазме (рис. 97). Ось, проходящая от анимального полю-
са к вегетативному, называется анимально-вегетативной осью. Количество желтка в олиголецитальных и мезолецитальных яйцеклетках возрастает по направлению от анимального к вегетативному полюсу.
Особенности дробления зиготы зависят от количества желтка и характера его распределения в цитоплазме зиготы (яйцеклетки). Различают два типа дробления, зависящие от количества желтка в яйце: 1) полное, или голобластическое дробление, свойственное зиготам, образующимся из гомолецитальных и мезолецитальных яиц; 2) неполное, или меробластическое дробление, характерное для зигот, образующихся из яиц, содержащих большое количество желтка (полилецитальные и мезолецитальные яйца), который при дроблении не делится. Полное (голобластическое) дробление (рис. 98) бывает равномерным (ланцетник) и неравномерным (амфибии). У последних образуются малые и большие бластомеры, называемые соответственно микромерами и макромерами (рис. 99).
В зависимости от количества содержащегося желтка яйцеклетки (яйца) подразделяются следующим образом.
Яйцеклетки (яйца)
По расположению желтка в цитоплазме выделяют следующие типы яйцеклеток.
Яйцеклетки (яйца)
Желток распределён равномерно
Желток частично смещён к вегетативному полюсу
желток переполняет всё вегетативное и частично анимальное полушария
Желток локализован в центре яйцеклетки
Расположение образующихся из зиготы бластомеров в пространстве друг относительно друга послужило основанием для определения типов дробления, исходя из расположения бластомеров. Выделяют радиальное (ланцетник), спиральное (моллюски), билатеральное, или двустороннесимметричное (круглые черви), бисимметричное, или двусимметричное (гребневики) и анархичное (плоские черви) дробления .
При радиальном дроблении борозды деления (митотические веретена) ориентированы параллельно или перпендикулярно анимально-вегетативной оси яйцеклетки (рис. 98). Через такую бластулу проходит несколько плоскостей (осей) симметрии.
Спиральное дробление отличается нарушением такого соответствия (борозды деления располагаются наклонно к анимально-вегетативной
оси), и дочерние бластомеры располагаются как бы по спирали. Образующаяся при спиральном дроблении бластула (стерробластула) не имеет ни полости, ни даже одной плоскости симметрии (рис. 100).
Билатеральное дробление характеризуется наличием в формирующейся бластуле одной оси (плоскости) симметрии (рис. 101).
При бисимметричном дроблении формирующаяся бластула имеет две оси (плоскости) симметрии (рис. 101).
Анархичное деление резко выделяется от описанных выше неупорядоченным расположением бластомеров и отсутствием оси (плоскости) симметрии (рис. 102).
Особенности локализации желтка в полилецитальных яйцеклетках существенно влияют на форму и расположение группы делящихся клеток. Это послужило основанием для выделения дискоидального (рыбы, рептилии, птицы) и поверхностного (насекомые) дробления.
Стадия дробления разделяется на 2 фазы: 1) фаза синхронного дробления -характеризуется одинаковой для всех клеток скоростью деления, обеспечивающей синхронность их деления; 2) фаза бластуляции, на которой исчезает синхронность деления клеток (у млекопитающих 1-я фаза отсутствует).
Все указанные особенности строения яйцеклеток, типов и конкретных способов дробления определяют форму и строение бластулы. Различают:
1) целобластулу - однослойный пузырёк с большим бластоцелем (ланцетник); стенка целобластулы (бластодерма) имеет примерно одинаковую толщину и состоит из одного слоя бластомеров, незначительно различающихся диаметром; нижняя часть бластодермы, находящаяся вокруг вегетативного полюса, образует дно бластулы; верхняя часть бластодермы, окружающая анимальный полюс, получила название крыши (рис. 98); в бластуле некоторых кишечнополостных бластомеры заполняют весь её объём; такая бластула, названная морулой, не имеет бластоцеля;
2) амфибластулу, состоящую из мелких (микромеров) и крупных (макромеров) бластомеров (земноводные); относительно небольшой бластоцель смещён к анимальному полюсу так, что место его расположения ограничивается анимальным полушарием (рис. 99);
3) дискобластулу, напоминающую формой диск, расположенный вокруг анимального полюса (рис. 103) и отделённый от желтка бластоцелем в виде щели (пресмыкающиеся, птицы);
4) перибластулу, однослойная бластодерма которой окружает желток, плотно прилегая к его поверхности (насекомые); бластоцель в перибластуле отсутствует (рис. 104).
Дробление характеризуется увеличением в бластомерах количества ДНК по отношению к количеству цитоплазмы, а также ведущей ролью цитоплазмы бластомеров в их судьбе. Бластомерами унаследуются разные участки цитоплазмы зиготы, которые по-разному влияют на активацию генов. Поскольку цитоплазма унаследована только от яйцеклетки, то развитие на стадии дробления протекает как бы по материнскому пути.
Перетяжки, разделяющие дробящуюся яйцеклетку на все более мелкие клетки (бластомеры), называются бороздами дробления. Дробление это многократные митотические деления зиготы, в результате которых зародыш становится многоклеточным, не меняя при этом существенно своего объема. Образование многоклеточности первая и основная биологическая роль дробления. Вторая роль состоит в восстановлении ядерно-плазматического отношения, которое падает в ходе стадии большого роста ооцита. Видовые особенности процесса дробления определяются двумя основными параметрами: количеством и распределением желточных белков в цитоплазме (желток подавляет дробление); присутствием в цитоплазме факторов, которые влияют на ориентацию митотического веретена и время его образования. Дробление начинается вскоре после оплодотворения и заканчивается, когда у зародыша достигается новое равновесие между ядром и цитоплазмой. Дробление строго координированный процесс, находящийся под генетическим контролем.
В большинстве других случаев клеточной пролиферации в период между митозами происходит рост клеток. Клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы. В период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Это деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения G1-периода в интерфазе, тогда как митозы следуют друг за другом с большой скоростью.
Темпы увеличения числа клеток в период дробления намного выше, чем на стадии гаструляции. Одним из последствий высокой интенсивности делений в процессе дробления является постепенное уменьшение отношения объема цитоплазмы к объему ядра. Изменение скорости, с которой происходит снижение отношения объема цитоплазмы к объему ядра, у многих типов зародышей является решающим фактором, определяющим время активации некоторых генов. От митотического деления соматических клеток дробление отличается тем, что полученные в результате дробления клетки не растут, а поэтому с каждым следующим делением становятся всё более мелкими, при этом увеличивается только их количество, а зародыш в целом не растет. Получающиеся при дроблении клетки мало дифференцированы и сравнительно однородны.
Дробление (сегментация) у отдельных представителей разряда позвоночных имеет в общем одинаковое течение; однако, как уже было упомянуто выше, оно находилось под влиянием факторов, которые во время филогенеза воздействовали на развитие в виде последствий влияния внутренней и внешней среды, в которой организмы проживали во время своего родового развития (ценогенетические факторы).
При наблюдении за изменениями , происходящими в яйцеклетках согласно филогенетическому развитию яиц отдельных представителей разряда позвоночных, можно заметить, что яйцевые клетки в значительной мере отличаются друг от друга по содержанию питательного и строительного вещества - желтка. Яйцевые клетки ланцетника (Amfioxus), организма, который в филогенетическом отношении считается наиболее низко организованным существом, но который уже обладает прочной спинной областью, относятся к числу олиголецитальных.
Однако, в соответствии с филогенетическим развитием , количество желтка в яйцеклетках позвоночных животных, являющихся филогенетически наиболее высоко организованными организмами, все более возрастает, достигая максимального количества в птичьих яйцеклетках, которые являются относительно очень крупными и полилецитальными. Под влиянием ценогенетических факторов (факторов, воздействующих из внешней среды и обусловливаемых изменением образа жизни, а следовательно, и развития) количество желтка в процессе филогенетического развития по направлению к человеку все более уменьшается, благодаря чему яйцеклетки человека и высших млекопитающих становятся снова (вторично) олиголецитальными.
Наличие вариабельного количества желтка
оказывает, как уже было сказано выше, значительное влияние на процесс дробления яйцеклетки. Яйцевые клетки с малым содержанием желтка (олиголецитальные) дробятся полностью, то есть все вещество оплодотворенного яйца при дроблении делится на новые клетки, бластомеры (яйцеклетки голобластического вида). Наоборот, у яйцеклеток, содержащих желтка больше, или даже большое количество желтка (полилецитальных), борозды дробления непрерывно дробят только меньшую часть ооплазмы, расположенную на так называемом анимальном полюсе, где желточных гранул меньше (яйцеклетки меробластического вида).
В соответствии с этим у отдельных представителей разряда
позвоночных различаются следующие типы дробления.
1. Полное дробление . К полному, тотальному дроблению относятся те случаи, когда в процессе дробящего деления делится вся оплодотворенная яйцевая клетка и борозды дробления распространяются по всей ее поверхности. По этому типу дробятся яйцевые клетки голобластического вида. В зависимости от содержания в ооплазме большего или меньшего количества желтка, а также в зависимости от его распределения в ооплазме, при дроблении возникают бластомеры либо сравнительно одинаковой величины (полное равномерное, эквальное, или адэквальное дробление), либо бластомеры различной величины, а именно более крупные в области с большим содержанием желтка и менее крупные в том месте, где желтка меньше (полное неравномерное, инэквальное дробление). Более крупные бластомеры называются макромерами, менее крупные - микромерами.
Полное эквальное, или адэквальное , дробление свойственно олиголецитальным, изолецитальным яйцеклеткам (ланцетник, высшие млекопитающие и человек); по полному инэквальному типу дробятся мезолецитальные яйцевые клетки анизолецитального и умеренно телолецитального вида (некоторые низшие рыбы и земноводные).
2. Частичное, парциальное, дробление . По частичному типу дробятся яйцевые клетки, содержащие значительное количество желтка (полилецитальные яйцеклетки), у которых из-за их больших размеров борозды дробления при клеточном делении проникают только в область анимального полюса, где находится клеточное ядро и где слой ооплазмы содержит меньше желточных гранул (высшие рыбы, пресмыкающиеся, птицы и некоторые низшие млекопитающие, яйцеродные).
При таком дроблении на анимальном полюсе сравнительно крупного яйца дробится только круглое поле (диск), в то время как остаток яйцевой клетки (желточный шар) остается не раздробленным (парциальное дисковидное дробление). У насекомых их полилецитальные центролецитальные яйцеклетки хотя и дробятся по всей поверхности, но центр клетки, содержащий большое количество желтка, остается не раздробленным (парциальное поверхностное дробление).
Строение бластулы
Дробление приводит к образованию шарообразного зародыша – бластулы . Если образуется сплошной шар без полости внутри, то такой зародыш называют морулой. Образование бластулы или морулы зависит от свойств цитоплазмы. Бластула образуется при достаточной вязкости цитоплазмы, морула – при слабой вязкости. При достаточной вязкости цитоплазмы бластомеры сохраняют округлую форму и только в местах соприкосновения слегка сплющиваются. Вследствие этого между ними появляется щель, которая по мере дробления увеличивается, заполняется жидкостью и превращается в бластоцель. При слабой вязкости цитоплазмы бластомеры не округляются и располагаются тесно друг возле друга, щели нет и полость не образуется. Бластулы различны по своему строению и зависят от типа дробления.
Типы бластул
Различают пять типов бластул : целобластулу, амфибластулу, стерробластулу, дискобластулу и перибластулу. Целобластула образуется при полном равномерном дроблении из яйцеклеток гомолецитального типа (ланцетник). Бластодерма целобластулы состоит из одного ряда более или менее одинаковых бластомеров, внутри находится крупная полость – бластоцель.
Бластодерма амфибластулы состоит из нескольких рядов клеток. Бластодерма в анимальной части тоньше, чем в вегетативной. Бластоцель меньших размеров, чем у ланцетника, и смещена к анимальному полюсу. Такого типа бластула образуется при полном неравномерном дроблении и характерна для круглоротых и земноводных.
Стерробластула состоит из одного ряда крупных бластомеров, которые глубоко заходят в полость бластулы, бластоцель в связи с этим или очень малая, или отсутствует (некоторые членистоногие).
Дискобластула образуется при неполном дискоидальном дроблении. Бластоцель в виде узкой щели находится между зародышевым диском и желтком. Крыша бластулы представлена бластодермой, а дно желтком. Такая бластула характерна для костистых рыб, пресмыкающихся и птиц. Бластодерма перибластулы состоит из одного ряда клеток, которые окружают желток. Полость в ней отсутствует. Перибластула наблюдается у некоторых насекомых.